|
El potencial de reposo de una célula es producido por diferencias en la concentración de iones dentro y fuera de la célula y por diferencias en la permeabilidad de la membrana celular a los diferentes iones. El potencial de equilibrio de Nernst relaciona la diferencia de potencial a ambos lados de una membrana biológica en el equilibrio con las características relacionadas con los iones del medio externo e interno y de la propia membrana. Producto destacado
Un caso prácticoImaginémonos un medio con KCl (cloruro de potasio), una sal, que, en medio acuoso, se disocia en K+y Cl-, sus iones componentes, que en la disolución se mantendrán en relación 1:1, compensando las cargas positivas de los cationes potasio con las negativas de los aniones cloruro, por lo que la disolución será electricamente neutra. Ahora añadamos a este modelo una membrana biológica selectivamente permeable, que permita el paso únicamente al K+. Así, los K+ difundirán libremente a un lado y otro de la membrana. Sin embargo, como hay más más iones en el compartimento 1, inicialmente fluirán más iones K+ del 1 al 2 que del 2 al 1. Como el Cl- no puede difundir a través de la membrana, pronto hay un exceso de carga positiva en el compartimento 2 y un exceso de carga negativa en el 1. Estas cargas en exceso se concentran a lo largo de la membrana, mientras que la masa de fluido en cada compartimento permanece neutra. Las capas de carga positiva y negativa a cada lado de la memrana producen una diferencia de potencial V = V1 − V2 a través de la membrana y un campo eléctrico E, que retarda el flujo de iones positivos del compartimento 1 al 2 y que acelera su flujo del compartimento 2 al 1. Tras un tiempo se alcanzará el equilibrio dinámico en el que exista un flujo de K+ idéntico del 2 al 1 como del 1 al 2. Este equilibrio depende de:
Además, la diferencia de potencial V en el equilibrio viene dada, en función de las concentraciones c1 y c2 de los iones de K+ en los dos compartimentos, mediante: Éste es el potencial de equilibrio de Nernst. Es negativo cuando la membrana es permeable a los iones positivos, y positivo cuando lo es a los aniones. Aquí k es la constante de Boltzmann y la T la temperatura absoluta. La magnitud kT es proporcional a la energía cinética media de los iones en solución y Modificaciones: Temperatura de 37 ºCA la temperatura corporal de 37 ºC el valor de de modo que el potencial de Nernst es Potencial de la membrana plasmáticaLa membrana plasmática de una célula nerviosa separa un citoplasma con una concentración de iones K + de 0,141 mol/L de un fluido extracelulular de únicamente 0,005 mol/L del mismo ion. Considerando que ambas concentraciones estén en equilibrio, y aplicando la fórmula anterior podemos deducir el potencial de membrana en reposo: donde este -89,5 mV teórico coincide aproximadamente con las mediciones en reposo efectuadas por técnicas de medición por microelectrodos en células vivas, de lo cual se deduce que las concentraciones de K + están casi en equilibrio. Sin embargo, para el mantenimiento de estas características se requiere una bomba iónica que propulse de forma dependiente de energía los cationes K + al interior celular. Transmisión del impulso nerviosoAdemás, los procesos de despolarización e hiperpolarización de la membrana durante, por ejemplo, la transmisión del impulso nervioso por parte de los potenciales de acción o la contracción muscular implican a un gran número de proteínas de membrana que intervienen en el flujo iónico bidireccional. De ahí que varíe tanto el potencial de membrana en estos casos: por ejemplo, una neurona pude despolarizarse hasta 30 mV, incrementando el potencial en más de 120 mV. Véase tambiénReferencias
|
|
| Este articulo se basa en el articulo Potencial_de_Nernst publicado en la enciclopedia libre de Wikipedia. El contenido está disponible bajo los términos de la Licencia de GNU Free Documentation License. Véase también en Wikipedia para obtener una lista de autores. |


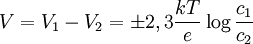
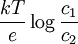 es proporcional al flujo neto de iones debido a la diferencia de concentración. La magnitud eV es proporcional al flujo neto de iones debido a la diferencia de potencial. Así, la ecuación antes citada es la condición para que estos dos flujos sean iguales y opuestos.
es proporcional al flujo neto de iones debido a la diferencia de concentración. La magnitud eV es proporcional al flujo neto de iones debido a la diferencia de potencial. Así, la ecuación antes citada es la condición para que estos dos flujos sean iguales y opuestos.
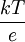 es:
es:
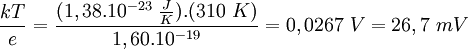
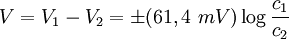
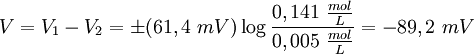
Último visto


